
奀憯晅拝嵶嬠偵傛傞娐嫬儌僯僞儕儞僌
孎杮戝妛増娸堟娐嫬壢妛嫵堢尋媶僙儞僞乕
嫵庼丂戨旜丂恑
暯惉24擭10寧24擔乮悈乯
侾丏偼偠傔偵
梴怋僲儕偼桳柧奀丒敧戙奀偵偍偗傞廳梫側悈嶻帒尮偺傂偲偮偱偡偑丄暯惉12擭偵偼怺崗側乬怓棊偪乭栤戣側偳偱惗嶻検偼寖尭偟丄幮夛栤戣偲側傝傑偟偨丅巹偼丆暯惉13擭偺増娸堟娐嫬壢妛嫵堢尋媶僙儞僞乕愝抲傪婡偵僲儕偵偮偄偰婎慴尋媶傪奐巒偟丆尰嵼傑偱乽僗僒價僲儕乮梴怋僲儕乯怓棊偪偺暘巕婡峔乿偲乽僗僒價僲儕偺儗僩儘僩儔儞僗億僝儞乿傪拞怱偵尋媶傪恑傔偰偒傑偟偨乮恾侾乯丅
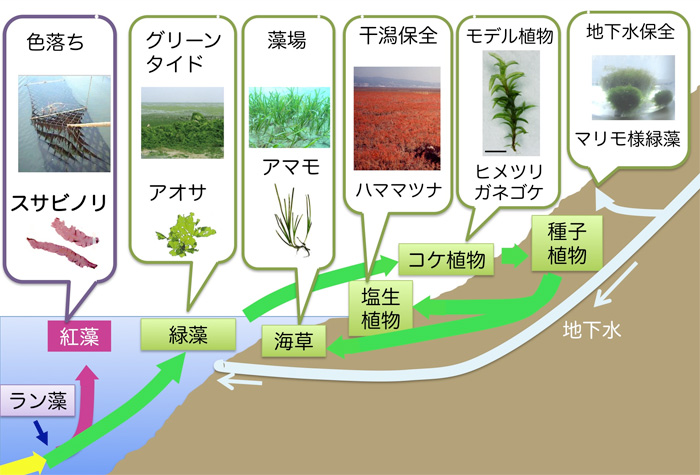
恾侾丂尋媶僥乕儅偲怉暔嵽椏
偦偺屻丄堩尒嫵庼偲偺嫟摨尋媶偲偟偰丄姳妰偺娐嫬曐慡偵廳梫側墫惗怉暔偺偆偪丄悽奅拞偵暘晍偡傞僴儅儅僣僫偺堚揱揑懡條惈夝愅傕巒傔傑偟偨丅偝傜偵丄奺抧偱戝検敪惗偑栤戣偵側偭偰偄傞傾僆僒偵偮偄偰傕堩尒嫵庼偲偺嫟摨尋媶傪峴偭偰偄傑偡丅僴儅儅僣僫傗傾僆僒偺尋媶偵偮偄偰偼梊旛揑抜奒偱偡偑丄偙偺巗柉島嵗傗増娸堟僙儞僞乕島墘夛偱傕偛徯夘偟偰偒傑偟偨丅乮孎杮戝妛増娸堟娐嫬壢妛嫵堢尋媶僙儞僞乕儂乕儉儁乕僕http://engan.kumamoto-u.ac.jp/index.html偺乽巗柉岞奐島嵗乿傗乽増娸堟僙儞僞乕島墘夛乿偺儁乕僕傪偛棗偔偩偝偄丅乯偝傜偵丄嶐擭偐傜丄敧戙奀嵞惗僾儘僕僃僋僩偲偟偰憯応奀憪傾儅儌偺晅拝嵶嬠偺尋媶傕巒傔偰偄傑偡丅傑偨丄抧壓悈傪拞怱偲偟偨悈娐嫬壢妛偵偮偄偰傕娭傢傞偙偲偵側傝乮http://www.gelk.info/乯丄僗僀僛儞僕僲儕梴怋応偱敪惗偡傞儅儕儌條憯椶偵偮偄偰傕挷傋偰偄傑偡丅崱夞偼丄埲慜偐傜栤戣偲側偭偰偄偨僗僒價僲儕偺攟梴僩儔僽儖偵偮偄偰丄堦嶐擭枛偵僩儔僽儖偺尒傜傟偨攟梴塼偐傜怴庬偺嵶嬠偑暘棧偝傟偨偙偲偐傜傗偭偲侾偮偺恑揥偑摼傜傟丄偦偺惉壥傪嶐擭偺巗柉岞奐島嵗偱乽奀憯晅拝嵶嬠偵傛傞娐嫬儌僯僞儕儞僌乿偲戣偟偰偛徯夘偝偣偰偄偨偩偒傑偟偨丅崱夞偼丄偙偺怴庬偺嵶嬠偺惈幙偵偮偄偰偺尋媶惉壥偲僗僒價僲儕帺恎偑嶌傞峈嬠暔幙偵娭偡傞尋媶傪偛徯夘偟傑偡丅
俀丏増娸堟偺娐嫬栤戣
乽奀戂偺怓棊偪乿偼丄愒挭僾儔儞僋僩儞偺敪惗側偳偵傛傝奀悈拞偺塰梴拏慺擹搙偑掅壓偟丄奀戂偺怓慺偑暘夝偡傞偙偲偵傛傝婲偙傞偙偲偑柧傜偐偵側偭偰偄傑偡丅増娸堟偺娐嫬栤戣偲偟偰愒挭(red tide)傗憯応偺徚幐偼椙偔抦傜傟偰偄傑偡偑丄嵟嬤偱偼傾僆僒椶偑戝検敪惗偡傞僌儕乕儞僞僀僪乮green tide乯傕戝偒側栤戣偲側偭偰偄傑偡(恾俀)丅僌儕乕儞僞僀僪偼擔杮奺抧偱傒傜傟丄嬨廈偵偍偄偰傕攷懡榩側偳偼椙偔抦傜傟偰偄傑偡丅孎杮導偱偼懠導傎偳戝偒側栤戣偲偼側偭偰偄傑偣傫偱偟偨偑丄嵟嬤偼尠嵼壔偟丄怴偨側娐嫬栤戣偲偟偰杮奿揑偵庢傝慻傓昁梫偑偱偰偒傑偟偨丅憯応奀憯丄僲儕偺怓棊偪丄僌儕乕儞僞僀僪偄偢傟傕奀憯傗奀憪側偳偺怉暔偑増娸堟偺娐嫬僗僩儗僗偵懳偡傞墳摎偲峫偊傞偙偲偑偱偒傑偡丅 |
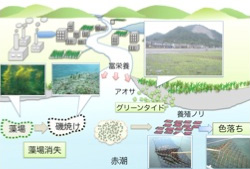 恾俀丂増娸堟偺娐嫬栤戣 |
俁丏僗僒價僲儕偺堦惗
怘梡偲偟偰偄傞僲儕偺梩忬懱偼敿悢悽戙偱偁傝丄搤婜偵惉挿偟丄弔偵側傞偲庴惛偟攞悢悽戙偺巺忬懱偲側傝傑偡丅巺忬懱偼奓妅偵怤擖偟偰壞婜傪夁偛偟傑偡乮恾3乯丅僗僒價僲儕偱偼僼儔僗僐撪偱攟梴偱偒傞弮宯偺攟梴姅偑暘棧偝傟偰偍傝丄巹偨偪偼偦傟傜傪梡偄偰尋媶傪峴偭偰偄傑偡丅巹払偼丄扨朎巕偐傜惗偠偨挿偝1乣5cm偺庒偄梩忬懱傪幚尡嵽椏偲偟偰巊梡偟偰偄傑偡丅梩忬懱偼15亷丄10帪娫柧婜/14帪娫埫婜丄嬻婥偺捠婥偵傛傝攟梴傪峴偭偰偄傑偡偑丄偙偺攟梴忦審偱偼梩忬懱偐傜偺惗怋婍姱偺宍惉偼尒傜傟傑偣傫乮恾係乯丅堦曽丄巺忬懱偺攟梴壏搙偼梩忬懱傛傝傕崅壏偺23亷偱丄楢懕柧強偱丄捠婥偼偍偙側傢偢惷抲攟梴偝傟偰偄傑偡丅
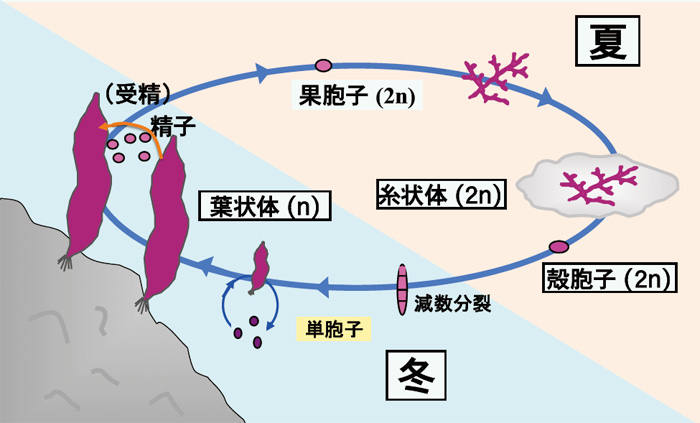 恾俁僗僒價僲儕偺惗妶娐 |
 恾係 僗僒價僲儕偺攟梴姅 |
係丏僗僒價僲儕偺婗敪惈僴儘僎儞壔桳婡壔崌暔崌惉
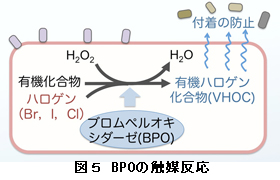 帺慠娐嫬壓偱偼奀憯昞柺偵柍悢偺嵶嬠偑晅拝偟偰偄傑偡偑丄奀憯偼桳奞側嵶嬠偺晅拝傪杊偖昁梫偑偁傞偱偟傚偆丅奀悈拞偵偼廘慺丄儓僂慺丄墫慺側偳偺僴儘僎儞椶偑懡検偵偁傞偨傔丄奀憯椶偼偙傟傜傪桳婡壔崌暔偲寢崌偝偣偨婗敪惈僴儘僎儞壔桳婡壔崌暔乮VHOC乯傪崌惉偟丄偦傟傪帺屓杊屼暔幙偲偟偰暘斿偟偰偄傞偙偲偑抦傜傟偰偄傑偡丅偙偺桳婡暔壔崌暔偵僴儘僎儞傪晅壛偡傞斀墳傪怗攠偡傞峺慺偑僽儘儉儁儖僆僉僔僟乕僛乮BPO乯偱偡乮恾俆乯丅惗堢抧偐傜嵦庢偝傟偨懡偔偺奀憯偼幚尡幒撪偱BPO妶惈傗VHOC惗惉擻傪傕偮偙偲偑曬崘偝傟偰偄傑偡丅堦曽丄傾儅僲儕懏偼BPO妶惈傕VHOC惗惉傕柍偄偙偲偑曬崘偝傟偰偄傑偟偨丅VHOC偼堦庬偺徚撆暔幙偱偡偺偱丄怘梡偲側傞傾儅僲儕懏偵偼VHOC崌惉擻偑側偄偙偲偼摉慠偺偙偲偺傛偆偵巚偊傑偟偨丅偟偐偟丄巹払偼丄俆擭慜偵幚尡幒撪偱攟梴偟偰偄傞巺忬懱偼丄BPO堚揱巕偑忢帪崅敪尰偟偰偄傞偑丄梩忬懱偼敪尰偑梷惂偝傟偰偄傞偙偲傪抦傝傑偟偨丅偦偟偰嵟嬤丄巺忬懱偼僽儘儌儂儖儉傪崌惉偡傞偙偲偑柧傜偐偵側傝傑偟偨丅偨偩偟丄埲慜偐傜巹払偺尋媶幒偱巊梡偟偰偄偨恖岺奀悈偲岝忦審偱偼VHOC偺惗惉偼傒傜傟傑偣傫偑丄攟梴塼偵僶僫僕僂儉偲僽儘儉傪揧壛偟丄岝嫮搙傪崅偔偡傞偲敪惗偟傑偟偨丅偦傟傜偺忦審偼帺慠娐嫬壓偺斖埻撪偲悇掕偝傟傑偟偨丅堦曽丄梩忬懱偼偙傟傜偺攟梴忦審偱偼VHOC傪惗惉偟傑偣傫丅偟偐偟丄廳嬥懏張棟側偳僗僩儗僗張棟傪峴偆偲BPO堚揱巕偺敪尰偑憹戝偟傑偡丅崱屻偼僗僩儗僗張棟偝傟偨梩忬懱偼VHOC傪敪惗偡傞偺偐柧傜偐偵偡傞梊掕偱偡丅
帺慠娐嫬壓偱偼奀憯昞柺偵柍悢偺嵶嬠偑晅拝偟偰偄傑偡偑丄奀憯偼桳奞側嵶嬠偺晅拝傪杊偖昁梫偑偁傞偱偟傚偆丅奀悈拞偵偼廘慺丄儓僂慺丄墫慺側偳偺僴儘僎儞椶偑懡検偵偁傞偨傔丄奀憯椶偼偙傟傜傪桳婡壔崌暔偲寢崌偝偣偨婗敪惈僴儘僎儞壔桳婡壔崌暔乮VHOC乯傪崌惉偟丄偦傟傪帺屓杊屼暔幙偲偟偰暘斿偟偰偄傞偙偲偑抦傜傟偰偄傑偡丅偙偺桳婡暔壔崌暔偵僴儘僎儞傪晅壛偡傞斀墳傪怗攠偡傞峺慺偑僽儘儉儁儖僆僉僔僟乕僛乮BPO乯偱偡乮恾俆乯丅惗堢抧偐傜嵦庢偝傟偨懡偔偺奀憯偼幚尡幒撪偱BPO妶惈傗VHOC惗惉擻傪傕偮偙偲偑曬崘偝傟偰偄傑偡丅堦曽丄傾儅僲儕懏偼BPO妶惈傕VHOC惗惉傕柍偄偙偲偑曬崘偝傟偰偄傑偟偨丅VHOC偼堦庬偺徚撆暔幙偱偡偺偱丄怘梡偲側傞傾儅僲儕懏偵偼VHOC崌惉擻偑側偄偙偲偼摉慠偺偙偲偺傛偆偵巚偊傑偟偨丅偟偐偟丄巹払偼丄俆擭慜偵幚尡幒撪偱攟梴偟偰偄傞巺忬懱偼丄BPO堚揱巕偑忢帪崅敪尰偟偰偄傞偑丄梩忬懱偼敪尰偑梷惂偝傟偰偄傞偙偲傪抦傝傑偟偨丅偦偟偰嵟嬤丄巺忬懱偼僽儘儌儂儖儉傪崌惉偡傞偙偲偑柧傜偐偵側傝傑偟偨丅偨偩偟丄埲慜偐傜巹払偺尋媶幒偱巊梡偟偰偄偨恖岺奀悈偲岝忦審偱偼VHOC偺惗惉偼傒傜傟傑偣傫偑丄攟梴塼偵僶僫僕僂儉偲僽儘儉傪揧壛偟丄岝嫮搙傪崅偔偡傞偲敪惗偟傑偟偨丅偦傟傜偺忦審偼帺慠娐嫬壓偺斖埻撪偲悇掕偝傟傑偟偨丅堦曽丄梩忬懱偼偙傟傜偺攟梴忦審偱偼VHOC傪惗惉偟傑偣傫丅偟偐偟丄廳嬥懏張棟側偳僗僩儗僗張棟傪峴偆偲BPO堚揱巕偺敪尰偑憹戝偟傑偡丅崱屻偼僗僩儗僗張棟偝傟偨梩忬懱偼VHOC傪敪惗偡傞偺偐柧傜偐偵偡傞梊掕偱偡丅
俆丏僗僒價僲儕梩忬懱偺晅拝嵶嬠
僗僒價僲儕偺梩忬懱偼柍嬠攟梴偡傞偲僇儖僗忬偵側傝惓忢側梩忬懱偑宍惉偝傟側偄偙偲偑曬崘偝傟偰偄傑偡丅杒奀摴戝妛偺嵉夈捈湋嫵庼偺僌儖乕僾偼僗僒價僲儕梩忬懱傪惓忢側宍懺偵桿摫偡傞擻椡偺偁傞嵶嬠悢庬傪僗僒價僲儕梩忬懱偐傜2004擭偵暘棧偟偰偄傑偡(Mori et al.) 丅偦偺屻丆僗僒價僲儕梩忬懱偵偼懡悢偺嵶嬠偑晅拝偟偰偄傞偲峫偊傜傟偰偄傑偟偨偑丆晅拝嵶嬠偺栐梾揑暘棧偼峴傢傟偰偄傑偣傫偱偟偨丅偟偐偟丆2010擭偵僗僒價僲儕梩忬懱偐傜23庬偺嵶嬠16SrRNA堚揱巕攝楍偑暘棧偝傟傑偟偨乮Namba et al.,2010乯丅偙偺曬崘偱偼嵉夈尋媶幒偱暘棧偝傟偨梩忬懱宍惉傪懀恑偡傞嵶嬠偼暘棧偝傟偰偄傑偣傫偱偟偨丅偙傟傜偺偙偲偐傜丆僗僒價僲儕梩忬懱偵偼偙傟傜埲奜偺嵶嬠傕晅拝偟偰偄傞壜擻惈偑峫偊傜傟傑偟偨丅巹払傕丆惓忢側忬懺偱惗堢偟偰偄傞梩忬懱偐傜晅拝嵶嬠傪暘棧偟偨偲偙傠丆梊憐傛傝傕庬椶偼彮側偔係庬偺攝楍(BPyGA1, BPyGA2, BPyGA3, BPyGB5)偑暘棧偝傟傑偟偨丅偙傟傜偼丆懠偺尋媶僌儖乕僾偐傜曬崘偝傟偨嵶嬠偲偼堎側偭偰偄傑偟偨丅
俇丏僗僒價僲儕梩忬懱偺攟梴僩儔僽儖
 巹偨偪偼丄僗僒價僲儕偺梩忬懱傪恖岺婥徾婍撪偱攟梴偟偰偄傑偡丅埲慜偼恖岺婥徾婍傪楲壓偵偍偄偰巊梡偟偰偄傑偟偨偑丄俆嶐擭偺戝妛偺夵廋岺帠廔椆屻偼丄偦傟傜偼怴抸偺幚尡幒偵堏偝傟傑偟偨丅偟偐偟丄堏愝摉弶偐傜梩忬懱傪怴慛攟抧偵堏偡偲攟梴塼偑敀戺壔偟梩忬懱偺惉挿偑埆偔尋媶傪恑傔傞偙偲偑偱偒側偔側傝傑偟偨乮恾俇乯丅晄巚媍側偙偲偵丄摨偠幚尡幒撪偵偁傞僲儕偺巺忬懱傗僲儕埲奜偺怉暔偼惓忢偵惗堢偟偰偄傑偟偨丅偦偺屻丄僲儕梩忬懱偺惗挿慾奞偺尨場傪扵偭偰偄偔偆偪偵丄幚尡幒撪偺嬻婥偵婗敪惈桳婡壔崌暔(VOC)偑娷傑傟偰偄傞壜擻惈偑偁傞偙偲偑暘偐傝傑偟偨丅偦偙偱丄婔偮偐偺晹壆偱攟梴傪帋傒偨寢壥丄巹偑巊梡偟偰偄傞嫵堳尋媶幒偵堏偡偲徢忬偼姰慡偵徚幐偟偰丄埲慜偲摨條偵惓忢偵惗堢偡傞偙偲偑柧傜偐偵側傝傑偟偨丅偦偙偱丄巇曽側偔僲儕偺梩忬懱偩偗偼巹偺晹壆偱攟梴偟偰偄傑偡丅偦偺屻丄幒撪僈僗惉暘傪暘愅偟偨偲偙傠丄梩忬懱攟梴塼偑敀戺壔偡傞晹壆偵偼丄偛偔旝検側偑傜偑懠偺晹壆偱偼専弌偝傟側偄僕僋儘儘儊僞儞丄2-僄僠儖-1-僿僉僒僲乕儖丄僄僞僲乕儖偑専弌偝傟傑偟偨丅梩忬懱攟梴偱偼丄幒撪嬻婥傪僄傾乕億儞僾偱媧偄崬傒丄僼傿儖僞乕傪捠偟偰柍嬠嬻婥傪攟梴塼偵憲傝崬傒丄塼傪奾漚偟偰偄傑偡丅2-僄僠儖-1-僿僉僒僲乕儖偼悈偵偼傎偲傫偳梟偗側偄偙偲偐傜丄敀戺壔偺尨場暔幙偼僄僞僲乕儖偐僕僋儘儘儊僞儞偺壜擻惈偑峫偊傜傟傑偟偨丅
巹偨偪偼丄僗僒價僲儕偺梩忬懱傪恖岺婥徾婍撪偱攟梴偟偰偄傑偡丅埲慜偼恖岺婥徾婍傪楲壓偵偍偄偰巊梡偟偰偄傑偟偨偑丄俆嶐擭偺戝妛偺夵廋岺帠廔椆屻偼丄偦傟傜偼怴抸偺幚尡幒偵堏偝傟傑偟偨丅偟偐偟丄堏愝摉弶偐傜梩忬懱傪怴慛攟抧偵堏偡偲攟梴塼偑敀戺壔偟梩忬懱偺惉挿偑埆偔尋媶傪恑傔傞偙偲偑偱偒側偔側傝傑偟偨乮恾俇乯丅晄巚媍側偙偲偵丄摨偠幚尡幒撪偵偁傞僲儕偺巺忬懱傗僲儕埲奜偺怉暔偼惓忢偵惗堢偟偰偄傑偟偨丅偦偺屻丄僲儕梩忬懱偺惗挿慾奞偺尨場傪扵偭偰偄偔偆偪偵丄幚尡幒撪偺嬻婥偵婗敪惈桳婡壔崌暔(VOC)偑娷傑傟偰偄傞壜擻惈偑偁傞偙偲偑暘偐傝傑偟偨丅偦偙偱丄婔偮偐偺晹壆偱攟梴傪帋傒偨寢壥丄巹偑巊梡偟偰偄傞嫵堳尋媶幒偵堏偡偲徢忬偼姰慡偵徚幐偟偰丄埲慜偲摨條偵惓忢偵惗堢偡傞偙偲偑柧傜偐偵側傝傑偟偨丅偦偙偱丄巇曽側偔僲儕偺梩忬懱偩偗偼巹偺晹壆偱攟梴偟偰偄傑偡丅偦偺屻丄幒撪僈僗惉暘傪暘愅偟偨偲偙傠丄梩忬懱攟梴塼偑敀戺壔偡傞晹壆偵偼丄偛偔旝検側偑傜偑懠偺晹壆偱偼専弌偝傟側偄僕僋儘儘儊僞儞丄2-僄僠儖-1-僿僉僒僲乕儖丄僄僞僲乕儖偑専弌偝傟傑偟偨丅梩忬懱攟梴偱偼丄幒撪嬻婥傪僄傾乕億儞僾偱媧偄崬傒丄僼傿儖僞乕傪捠偟偰柍嬠嬻婥傪攟梴塼偵憲傝崬傒丄塼傪奾漚偟偰偄傑偡丅2-僄僠儖-1-僿僉僒僲乕儖偼悈偵偼傎偲傫偳梟偗側偄偙偲偐傜丄敀戺壔偺尨場暔幙偼僄僞僲乕儖偐僕僋儘儘儊僞儞偺壜擻惈偑峫偊傜傟傑偟偨丅
僄僞僲乕儖偑嶦嬠嵻偲偟偰棙梡偝傟偰偄傞傛偆偵丄僄僞僲乕儖帒壔惈嬠偼摿庩側嵶嬠偲尵偊傑偡丅僔儏乕僪儌僫僗偺堦庬Pseudomonas fluorescens strain S227偼僄僞僲乕儖傪扽慺尮偲偟偰惗堢偡傞帪偵丄峈僇價妶惈傪傕偮峈惗暔幙乮pyoluteorin側偳乯傪嶌傝傑偡丅偙偺嵶嬠偼丄怓乆側扽慺尮偺傕偲偱惉挿偱偒傑偡偑丄pyoluteorin傪偮偔傞偺偼僄僞僲乕儖偲僌儕僙儘乕儖偩偗偱偟偨P. fluorescens丂S227偼怉暔偺崻偵姶愼偟昦婥傪堷偒婲偡僇價椶偺惗暔杊屼嵻偲偟偰傛偔棙梡偝傟偰偄傑偡丅BPy亅侾偺僄僞僲乕儖戙幱嶻暔偑偳偺傛偆側惗棟婡擻傪傕偮偺偐嫽枴偁傞栤戣偱偡丅
俈.僗僒價僲儕梩忬懱偺晅拝嵶嬠
僗僒價僲儕梩忬懱偺攟梴偵傛傝惗偠偨敀戺攟梴塼偐傜嵶嬠傪堦庬乮BPy-侾乯暘棧偟丆16S儕儃僜乕儉RNA堚揱巕偺墫婎攝楍傪挷傋丆婛懚偺嵶嬠庬偺攝楍傪斾妑偟偨偲偙傠丆僯儏乕僕乕儔儞僪偺儈僪儕僀僈僀梴怋応偺僾儘僶僀僆僥傿僋偲偟偰2010擭偵暘棧偝傟偨僱僾僣僲儌僫僗懏(Neptunomonas)偺怴庬偺嵶嬠乮Kesarcodi-Watson et al. Aquaculture 309:49-55乯偲攝楍偑姰慡偵堦抳偟傑偟偨丅傑偨丆僌儔儉愼怓惈丆朎巕偺桳柍丆塣摦惈丆僇僞儔乕僛斀墳丆僆僉僔僟乕僛斀墳側偳偺惗壔妛僥僗僩偺寢壥傕忋婰偺嵶嬠偲傎傏堦抳偟傑偟偨丅師偵丆恖岺奀悈偵僕僋儘儘儊僞儞傗僄僞僲乕儖壛偊偰丆BPy-侾偺惉挿傪挷傋偨偲偙傠丆僕僋儘儘儊僞儞偱偼惉挿偑傒傜傟傑偣傫偱偟偨偑丆僄僞僲乕儖偱偼椙偔惉挿偡傞偙偲偑暘偐傝傑偟偨丅恖岺奀悈偵偼旝検偺價僞儈儞傪彍偄偰扽慺尮偼娷傑傟偰偄傑偣傫丅偙偺幚尡偐傜丆BPy-侾偼僄僞僲乕儖傪桞堦偺扽慺尮偲偟偰惉挿偱偒傞僄僞僲乕儖帒壔惈嬠偱偁傞偙偲偑傢偐傝傑偟偨丅
僄僞僲乕儖偑嶦嬠嵻偲偟偰棙梡偝傟偰偄傞傛偆偵丆僄僞僲乕儖帒壔惈嬠偼摿庩側嵶嬠偲尵偊傑偡丅僔儏乕僪儌僫僗偺堦庬Pseudomonas fluorescens strain S227偼僄僞僲乕儖傪扽慺尮偲偟偰惗堢偡傞帪偵丆峈僇價妶惈傪傕偮峈惗暔幙乮pyoluteorin側偳乯傪嶌傝傑偡丅偙偺嵶嬠偼丆怓乆側扽慺尮偺傕偲偱惉挿偱偒傑偡偑丆pyoluteorin傪偮偔傞偺偼僄僞僲乕儖偲僌儕僙儘乕儖偩偗偱偟偨P. fluorescens丂S227偼怉暔偺崻偵姶愼偟昦婥傪堷偒婲偡僇價椶偺惗暔杊屼嵻偲偟偰傛偔棙梡偝傟偰偄傑偡丅BPy-侾偺僄僞僲乕儖戙幱嶻暔偑偳偺傛偆側惗棟婡擻傪傕偮偺偐嫽枴偁傞栤戣偱偡丅
俉丏惓忢側梩忬懱攟梴塼偵僄僞僲乕儖傪揧壛偡傞偲敀戺壔偑桿摫偝傟傞
BPy亅侾偑僄僞僲乕儖帒壔惈嬠偱偁偭偨偙偲偐傜丄怉暔攟梴幒偱梩忬懱傪攟梴偟偨偲偒偵惗偠傞敀戺壔偼幒撪嬻婥偺僄僞僲乕儖偑攟梴塼偵梟夝偟BPy亅侾偺憹怋傪桿摫偟偨偺偱偼側偄偐偲峫偊傜傟傑偟偨丅偦偙偱丄惓忢側梩忬懱攟梴塼偵僄僞僲乕儖傪揧壛偟丄敀戺壔偑婲偙傜側偄嫵堳尋媶幒乮恾俇乯偱梩忬懱傪攟梴偟偨偲偙傠丄怉暔攟梴幒偺帪偲摨條偺敀戺壔偑尒傜傟傑偟偨丅偦偙偱丄僄僞僲乕儖偵傛偭偰惗偠偨敀戺塼偐傜嵶嬠DNA傪夞廂偟丄16SrRNA堚揱巕偺攝楍傪挷傋偨偲偙傠丄BPy亅侾偺懠偵傕偆堦庬椶偺嵶嬠BPyA俁偑暘棧偝傟傑偟偨丅偙傟偼丄僄僞僲乕儖柍揧壛偱惗偠偨梩忬懱敀戺塼偺応崌偲摨偠偱偟偨丅BPyA3偺16SrRNA堚揱巕偺墫婎攝楍偼BPy-1偲偼93亾堦抳偟偰偄傑偟偨丅BPyA3偺惗壔妛揑惈幙偼傑偩挷傋偰偄傑偣傫偑丄僄僞僲乕儖帒壔惈偱偁傞偙偲偼柧傜偐偱偡丅傑偨丄晄巚媍側偙偲偵僄僞僲乕儖揧壛偺応崌偵偍偄偰傕丄捠忢偺梩忬懱偵晅拝偟偰偄傞係庬偺嵶嬠乮BPyGA1, BPyGA2, BPyGA3, BPyGB5乯偺攝楍偼専弌偝傟傑偣傫偱偟偨丅
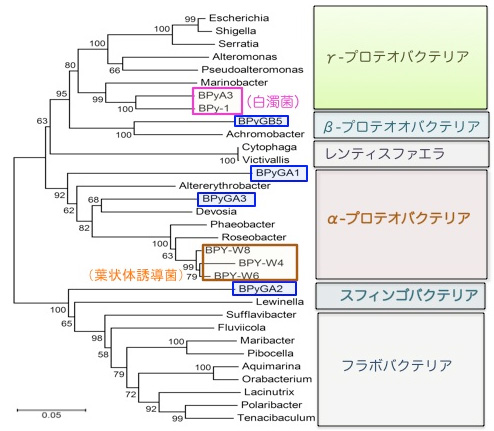
尰嵼傑偱偵僗僒價僲儕梩忬懱偐傜暘棧偝傟偨16SrRNA堚揱巕攝楍傪傕偲偵暘棧偝傟偨嵶嬠偺椶墢娭學傪暘巕宯摑庽偵傛傝恾帵偟偨偺偑恾7偱偡丅巹払偑暘棧偟偨嵶嬠俇庬偺攝楍偼懠偺尋媶僌儖乕僾偲偼堦抳偟傑偣傫偱偟偨丅偙傟偼梩忬懱偺宯摑姅偑堎側傞偙偲偵傛傞偲峫偊傜傟傑偡偑丄偦傟埲奜偵丄PCR偱巊梡偡傞僾儔僀儅乕偺堘偄偵傛傞壜擻惈傕巆偝傟偰偄傑偡丅
崱傑偱偺寢壥偐傜僗僒價僲儕梩忬懱偺敀戺壔傪師偺傛偆偵悇掕偟偰偄傑偡丅
侾乯僗僒價僲儕梩忬懱偵偼僄僞僲乕儖傪扽慺尮偲偟偰惗堢偱偒側偄係庬偺嵶嬠偑庡梫側嵶嬠偲偟偰晅拝偟偰偄傞丅俀乯俀庬偺僄僞僲乕儖帒壔惈嵶嬠傕懚嵼偡傞偑丄偙傟傜偼惓忢側攟梴忦審偱偼惉挿偼抶偔丄彫偝側廤抍偲偟偰堐帩偝傟偰偄傞丅俁乯僄僞僲乕儖傪娷傓幒撪偱攟梴偝傟傞偲丄僄僞僲乕儖帒壔惈嬠偼攟梴塼拞偺僄僞僲乕儖傪扽慺尮偲偟偰堎忢斏怋偡傞偨傔偵攟梴塼偑敀戺壔偡傞丅
巹偨偪偼丄攟梴塼偺敀戺壔偵傛偭偰僗僒價僲儕梩忬懱偼惗堢偑慾奞偝傟偰偄偨偨傔偵丄敀戺壔嵶嬠偼桳奞側嵶嬠偱偁傞偲峫偊偰偄傑偟偨丅偟偐偟丄BPy亅侾傗BPyA3偼桳奞暔幙偱偁傞僄僞僲乕儖傪彍嫀偡傞偨傔偺桳塿側嵶嬠偲偟偰曐帩偝傟偰偄傞偺偐傕偟傟傑偣傫丅
BPy亅侾傗BPyA3偑廬棃偺梩忬懱晅拝嵶嬠偺暘棧朄偱偼専弌偱偒側偐偭偨偙偲傪峫偊傞偲丄崱屻桳塿側嵶嬠傪暘棧偡傞偨傔偵偼丄捠忢偺嵶嬠暘棧朄偵壛偊偰僗僩儗僗偵墳摎偡傞嵶嬠傪暘棧偡傞偲偄偆曽朄偑桳岠偐傕偟傟傑偣傫丅崱夞偼僄僞僲乕儖偵傛偭偰桿摫偝傟傞嵶嬠傪尒偮偗傞偙偲偑偱偒傑偟偨偑丄崱屻偼丄偦偺懠偺僗僩儗僗張棟偵傛傝怴偨側嵶嬠偑暘棧偱偒傞偺偐挷傋傞梊掕偱偡丅
恾俈丂16SrDNA攝楍偵傛傞暘巕宯摑庽
俋丏僾儘僶僀僆僥傿僋僗偲梴怋嬈
僸僩偺挵撪偵偼堦恖摉偨傝100庬椶埲忋丄100挍屄埲忋偺挵撪嵶嬠偑惗懅偟偰偍傝丄偦傟傜偼僶儔儞僗傪曐偪側偑傜丄堦庬偺惗懺宯乮挵撪僼儘乕儔乯傪宍惉偟偰偄傞偲峫偊傜傟偰偄傑偡丅僾儘僶僀僆僥傿僋僗偲偼乽挵撪僼儘乕儔乮挵撪嬠憄乯偺僶儔儞僗傪夵慞偡傞偙偲偵傛傝恖偵桳塿側嶌梡傪傕偨傜偡惗偒偨旝惗暔乿偱偁傝丄擕巁嬠側偳偺桳梡嬠偼僾儘僶僀僆僥傿僋僗偲偟偰棙梡偝傟偰偄傑偡丅梴怋嫏嬈偱偼丄崅枾搙梴怋偵敽偆嫑昦敪惗偺梊杊丒帯椕偺偨傔偵栻嵻偑巊傢傟傞偙偲偑懡偄偱偡偑丄怘偺埨慡偺娤揰摍偐傜栤戣偑偁傝傑偡丅偨偲偊偽丄梴怋嫑偑姶愼徢傪敪徢偡傞偲峈惗暔幙偵傛傞壔妛椕朄偑偲傜傟傞偙偲偑懡偄偱偡偑丄峈惗暔幙偺懡梡偵傛傝栻嵻懴惈嬠偑敪惗偟偨傝丄廃曈悈堟偵棳弌偟偨峈惗暔幙偑娐嫬忩壔傪扴偭偰偄傞旝惗暔偺妶惈傪梷惂偡傞側偳偺栤戣偑婋湝偝傟傑偡丅偙偺懳墳嶔偲偟偰丄嫑椶徚壔娗傛傝暘棧偟偨擕巁嬠傪嫑偵搳梌偡傞偙偲偵傛偭偰丄偦偺僾儘僶僀僆僥傿僋僗岠壥偵傛傝丄埨慡側嫑傪堢偰傞尋媶偑峴傢傟偰偄傑偡丅偙偺傛偆偵僸僩傗嫑偼挵撪偵僾儘僶僀僆僥傿僋僗偲側傞嵶嬠傪傕偭偰偄傑偡丅僗僒價僲儕偺BPy亅侾偼僯儏乕僕乕儔儞僪偺儌僄僊僀僈僀梴怋応偺僾儘僶僀僆僥傿僢僋僗偲偟偰暘棧偝傟偨嵶嬠偲墫婎攝楍偑堦抳偡傞偙偲偐傜丄BPy亅侾偼僲儕埲奜偺惗暔偵傕桳梡側摥偒傪偡傞壜擻惈偑偁傝傑偡丅偝傜偵憐憸傪朿傜傑偣傞偲丄BPy亅侾偺晅拝偟偨僲儕傪梴怋偡傞偲丄奀戂梴怋応偺嬤偔偺懠偺嫑夘椶偺梴怋偵傕椙岲側岠壥傪梌偊傞壜擻惈偑偁傞偐傕偟傟傑偣傫丅