|
|
 |
| D.環境保全 > 2.環・阿蘇/有明・八代海の環境保全とブルー・グリーンツーリズム > 2-(1)「有明海・八代海を科学する」 |
| D.2-2-1 市民公開講座「有明海・八代海を科学する」
|
 |
|
平成16年10月21日
|
| 養殖ノリは有明海・八代海における重要な水産資源のひとつですが、平成12年には深刻な"色落ち"問題などで生産量は激減し、社会問題となりました。熊本大学は平成13年の沿岸域環境科学教育研究センター設置を機に、ノリについて基礎研究を開始しました。現在、スサビノリの遺伝子について熊本県水産研究センター、県内企業および他大学水産学部の協力のもとに研究を進めています。昨年度のこの講座では,「養殖ノリの色落ちのメカニズム」についての研究の一端を紹介しました。今回は,この研究の他に「スサビノリの動く遺伝子(レトロトランスポゾン)」についても紹介させていただきました。ここでは授業のうちで入門的解説の部分を掲載いたします。 | |
|
 「ノリの色落ち」を抑制する遺伝子 「ノリの色落ち」を抑制する遺伝子 |
| |
| 1.ノリとはどのような植物か? |
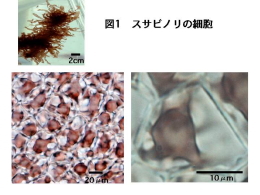
ノリ養殖に利用されているスサビノリやアサクサノリは紅藻類のアマノリ属(Porphyra)に属する海産の藻類です(図1はスサビノリの細胞の写真です。各細胞には紅色の葉緑体が一つ存在します)。
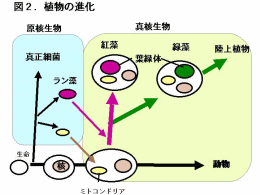
生物は原核生物と真核生物の二つに大別できます。原核生物では遺伝子の本体であるDNAが裸の状態で細胞内に存在していますが、真核生物ではDNAは核とよばれる膜に包まれた部位に閉じこめられています。真核生物はさらに植物と動物に大別することができ、植物ではDNAは核の他にミトコンドリアや葉緑体にも含まれています。植物の進化において、光合成を行う原核生物であるラン藻が植物の祖先となる細胞に取り込まれて、やがて細胞内で共生し現在の葉緑体になったと考えられています(一次共生、図2)。そして、葉緑体の性質などから紅藻は真核生物のなかでもっとも起源の古い植物であると考えられています。 |
|
|
| |
| 2.ノリの「色」のしくみとはたらき |
|
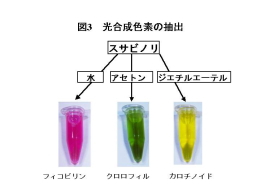
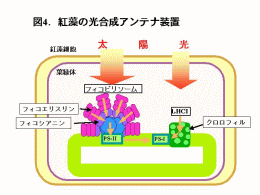
スサビノリを水や有機溶媒を用いて破砕するとフィコビリン、クロロフィル、カロチノイドなどの光合成色素が抽出できます(図3)。フィコビリンにはおもにフィコシアニン、フィコエリスリン、アロフィコシアニンが含まれています。これらは光合成の反応に必要な光を集めるためのアンテナの働きをする装置(フィコビリソーム)の構成色素です(図4)。緑藻や陸上植物はフィコビリンをもたず、アンテナ色素としてクロロフィルを使っているため、葉などの光合成器官は緑色をしています。光合成を行うバクテリアの一種であるラン藻やスサビノリの属する紅藻はアンテナ色素としてフィコビリンを使っている点で他の植物群とは異なっています。 |
|
|
| |
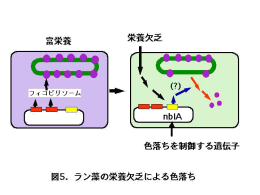
| 3.ラン藻の「色落ち誘導遺伝子」 |
ラン藻は窒素やリンなど栄養分が欠乏するとフィコビリソームを分解し、その産物
を栄養源として再利用することが知られています(図5)。ラン藻にとって「栄養欠
乏による色落ち」は過酷な環境で生き残るための重要な手段といえます。「色落ち能
を失った突然変異体」の遺伝子解析により、「色落ちを制御する遺伝子」は多数存在す
ることが明らかになってきました。そのうちでもっとも重要な遺伝子nblAは、通常の成育状態ではその働きが抑制されていますが、栄養欠乏になると働きはじめます。遺伝子組み換え技術を使ってラン藻のnblA遺伝子を破壊すると「色落ち能が失われる」ことも証明されています。
|
| |
| |
| 4.ノリの葉緑体DNAにもラン藻のnblAと類似の遺伝子がある |
養殖ノリもラン藻と同様に栄養欠乏になるとフィコビリンが減少することは、およそ15年前に詳しく研究されていました。それによると、フィコビリンの他に、クロロフィルやカロチノイドも減少しています。平成12年末の有明海で起こった色落ちノリも同じ症状を示していました。栄養欠乏によりフィコビリンが減少することから、ノリにもラン藻と同じようなしくみがあるのではないかと考えられました。
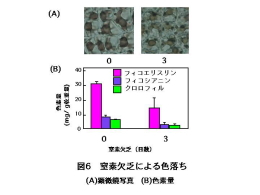
私たちはスサビノリを人工海水を用いて実験室内で培養し、栄養欠乏により色落ちノリを作成し、葉緑体や色素の変化を調べたところ、窒素欠乏培養液に移すとフィコビリンやクロロフィルの減少が観察できました(図6)。また,リン欠乏や鉄欠乏でもフィコビリンやクロロフィルの減少が観察できました。 |
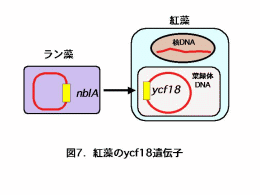
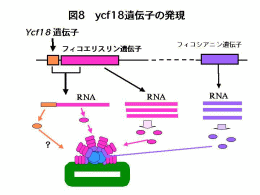
スサビノリと同じアマノリ属のパープレアアマノリでは葉緑体DNAの全塩基配列が決定されています。パープレアアマノリやその他の紅藻の葉緑体DNA中にもラン藻のnblAとよく似た遺伝子ycf18が存在していることは知られていましたが、その働きについては調べられていませんでした(図7)。そこで、パープレアアマノリの遺伝子情報を参考にして、スサビノリからycf18とその近傍の遺伝子断片を分離し塩基配列を決定しました。その遺伝子構造はパープレアアマノリと同じで、ycf18遺伝子はフィコエリスリン遺伝子と隣接して存在していました(図8)。 |
| |
| そして,ycf18遺伝子の発現状態を調べたところ、いくつかの点でラン藻のnblAとは異なることが分かりました。遺伝子はDNAの4種類の遺伝暗号として保存されています。遺伝子が働くためには,まず,遺伝暗号がRNAに読み取られます。そして,RNAの情報をもとにタンパク質が合成されます。ycf18遺伝子の働きを知るために,まず,ycf18のRNA量を調べました。通常の栄養状態のノリからRNAを分離してycf18のRNA量を調べたところ,微量ですが検出されました。また,ycf18は単独でRNAが作られるのではなく,フィコエリスリンRNAと一緒に合成されていました。また,窒素欠乏ではフィコエリスリンRNAだけてなくycf18RNAも見られなくなりました。このことは,ラン藻のnblAとは反対の反応でした。ycf18RNAが合成されるときには同時にフィコエリスリンRNAも作られることから,
ycf18が「色落ち(フィコビリソームの分解)」に働くのではなく,むしろ,「フィコビリソーム作り」に働いている可能性が生まれました。私達は,窒素欠乏以外の栄養条件を検討し,「
ycf18はフィコビリソームの再構築に働く」のではないかの考え研究を進めています。 |
|
|
| |
 「動く遺伝子(レトロトランスポゾン)」による新品種作出 「動く遺伝子(レトロトランスポゾン)」による新品種作出 |
|
|
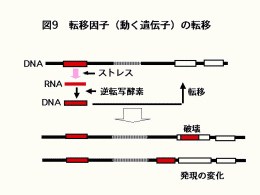
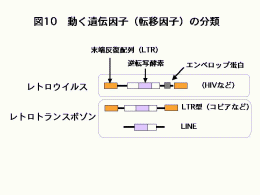
35億年の生命の歴史における革新的なできごとして,20億年前の真核生物の誕生と10億年前の多細胞生物の誕生が挙げられます。このとき細胞あたりのDNA量が大きく増大しています。このDNA量の増大の原因のひとつは遺伝子量の増大が考えられますが,もう一つのメカニズムとして動く遺伝子(転移因子)の働きも考えられています(図9)。「寄生体のようなDNA断片」とも呼ばれている転移因子は,含まれている配列によっていくつかに分類されています(図10)。DNA因子がRNAに読み取られ,そのRNAから逆転写酵素野働きでDNAが作られ,そのDNAが染色体の他の部位に挿入(転移)されるものはレトロトランスポゾンと呼ばれています。レトロトランスポゾンはLTRと呼ばれる末端反復配列をもつLTR型レトロトランスポゾンやLTRをもたないLINEなどに大別されます。真核生物のDNA中には様々なレトロトランスポゾンが存在しています。しかし,その多くは過去に動き廻り,今では動くことができなくなった残骸となっているものです。一方,幾つかのレトロトランスポゾンは動くことはできるが通常は活動を停止しているものもあります。
イネのTos17と呼ばれるLTR型レトロトランスポゾンはDNA中に1?数個しかありませんが,プロトプラスト処理や長期間の細胞培養により動き出し,DNA中に無数の子孫を挿入することが知られています。イネではプロトプラスト処理を利用して多くの変異体が作成されています。
| |
養殖ノリの場合にもプロトプラストからの再生植物には様々な形態変異が見られることが知られていました。私達はこの現象はレトロトランスポゾンの働きよるものではないかと考え,スサビノリの転移能をもつレトロトランスポゾンの分離をめざして研究を行っています。転移能をもつレトロトランスポゾンが分離できると,それらを利用して多くの変異体を作出することができると考えられます。
レトロトランスポゾンによる変異体作出は新品種の作出を容易にするだけではなく,遺伝子の機能を解析するうえでも有用です。スサビノリは実験生物としての利点をもつことから、大型海藻類の実験モデル生物として注目されており,その遺伝子解析にレトロトランスポゾンの研究は貢献できると考えられます。スサビノリは従来の食品としての価値だけではなく、遺伝子情報を利用した海藻類特有の新規有用物質の生産など新たな産業創製の可能性も秘めています。
|
|
| |
|
| 【熊本大学 地域貢献特別支援事業】 |