
養殖ノリの色落ちと環境ストレス応答
熊本大学沿岸域環境科学教育研究センター
教授 瀧尾 進
平成21年11月11日(水)
【1.はじめに】
養殖ノリは有明海・八代海における重要な水産資源のひとつですが、平成12年には深刻な“色落ち”問題などで生産量は激減し、社会問題となりました。私達は,平成13年の沿岸域環境科学教育研究センター設置を機にノリについて基礎研究を開始し,現在まで「スサビノリ(養殖ノリ)色落ちの分子機構」と「スサビノリのレトロトランスポゾン」を中心に研究を進めてきました。
今回は栄養欠乏と重金属処理という,異なる環境ストレスによって生じるノリの色落ちについての研究例を紹介し,色落ちの仕組みや働きについて植物の生理学や遺伝子科学の観点から解説致します。
【2.海苔養殖の歴史】
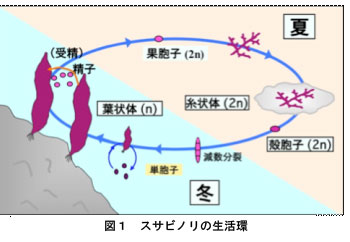 ノリ養殖に利用されているスサビノリやアサクサノリは紅藻類のアマノリ属(Porphyra)に属する海産の藻類です。海苔養殖では以前にはアサクサノリが利用されていましたが,スサビノリの方が生長が早く,病気にも強いことから,現在では養殖ノリのほぼすべてがスサビノリ系統となっています。一方,アサクサノリは現在では絶滅危惧種となっています。しかし,アサクサノリはスサビノリよりもが味が良いことからアサクサノリ養殖の復活も始まっています。図1はスサビノリの一生の模式図です。食用となる葉状体は半数世代(n,染色体数が2nの半分)で冬に生長します。春先になり精子と卵が葉状体に形成され,受精により受精卵(果胞子)ができ複数世代(2n)となります。果胞子が発芽すると葉状体とは全く異なる形態の糸状体となります。しかも,糸状体は貝殻の表面に付着し,その後,殻の中に侵入して殻の中でカビの菌糸のように増殖します。糸状体の生長は夏期に行われ,秋になると糸状体の一部から殻胞子(2n)が作られ,これが減数分裂した後に発芽し,半数世代の小さな葉状体となり,やがて大きな葉状体へと成長します。
ノリ養殖に利用されているスサビノリやアサクサノリは紅藻類のアマノリ属(Porphyra)に属する海産の藻類です。海苔養殖では以前にはアサクサノリが利用されていましたが,スサビノリの方が生長が早く,病気にも強いことから,現在では養殖ノリのほぼすべてがスサビノリ系統となっています。一方,アサクサノリは現在では絶滅危惧種となっています。しかし,アサクサノリはスサビノリよりもが味が良いことからアサクサノリ養殖の復活も始まっています。図1はスサビノリの一生の模式図です。食用となる葉状体は半数世代(n,染色体数が2nの半分)で冬に生長します。春先になり精子と卵が葉状体に形成され,受精により受精卵(果胞子)ができ複数世代(2n)となります。果胞子が発芽すると葉状体とは全く異なる形態の糸状体となります。しかも,糸状体は貝殻の表面に付着し,その後,殻の中に侵入して殻の中でカビの菌糸のように増殖します。糸状体の生長は夏期に行われ,秋になると糸状体の一部から殻胞子(2n)が作られ,これが減数分裂した後に発芽し,半数世代の小さな葉状体となり,やがて大きな葉状体へと成長します。
【3.ノリの植物学上の特徴】
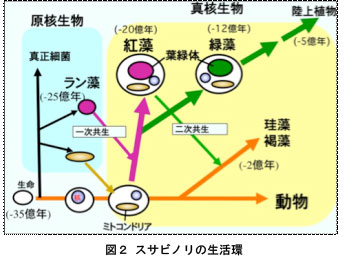 生物は原核生物(バクテリア)と真核生物の二つに大別できます(図2)。原核生物では遺伝子の本体であるDNAが裸の状態で細胞内に存在していますが、真核生物ではDNAは核とよばれる膜に包まれた部位に閉じこめられています。真核生物はさらに植物と動物に大別することができ、植物ではDNAは核の他にミトコンドリアや葉緑体にも含まれています。植物の進化において、光合成を行う原核生物であるラン藻が植物の祖先となる細胞に取り込まれて、やがて細胞内で共生し現在の葉緑体になったと考えられています(一次共生、図2)。すなわち,紅藻は植物進化において最初に出現した植物と言えます。紅藻が出現した後,緑藻が生まれ,その仲間は陸上へと進出しました。なお,コンブやワカメなどの褐藻類や赤潮プランクトンに含まれる珪藻や渦鞭毛藻類は,紅藻が動物細胞にもう一度取り込まれることによって生まれた(二次共生)植物と考えられています。このように、同じ海藻でも紅藻と褐藻は起原が大きく異なり,紅藻は真核生物のなかでもっとも起源の古い植物であることは,「ノリの色落ち」を考える上でも重要な点です。
生物は原核生物(バクテリア)と真核生物の二つに大別できます(図2)。原核生物では遺伝子の本体であるDNAが裸の状態で細胞内に存在していますが、真核生物ではDNAは核とよばれる膜に包まれた部位に閉じこめられています。真核生物はさらに植物と動物に大別することができ、植物ではDNAは核の他にミトコンドリアや葉緑体にも含まれています。植物の進化において、光合成を行う原核生物であるラン藻が植物の祖先となる細胞に取り込まれて、やがて細胞内で共生し現在の葉緑体になったと考えられています(一次共生、図2)。すなわち,紅藻は植物進化において最初に出現した植物と言えます。紅藻が出現した後,緑藻が生まれ,その仲間は陸上へと進出しました。なお,コンブやワカメなどの褐藻類や赤潮プランクトンに含まれる珪藻や渦鞭毛藻類は,紅藻が動物細胞にもう一度取り込まれることによって生まれた(二次共生)植物と考えられています。このように、同じ海藻でも紅藻と褐藻は起原が大きく異なり,紅藻は真核生物のなかでもっとも起源の古い植物であることは,「ノリの色落ち」を考える上でも重要な点です。
【4.ノリの色と葉緑体】
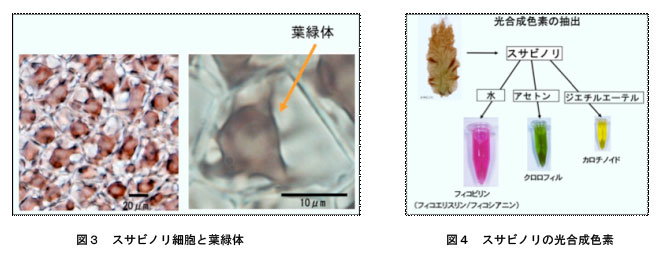
植物は外部環境の変化に対しさまざまな応答を示しますが,なかでも,エネルギー生産の場である葉緑体では反応中心複合体やアンテナ色素複合体は光条件だけではなく栄養欠乏などの各種ストレスにも応答しダイナミックな変動を示します。図3はスサビノリの葉状体の細胞の顕微鏡写真です(左は低倍率,右は高倍率で1つの細胞の中に金平糖状の葉緑体がある)。また,図4はスサビノリ葉状体を各種の溶液で破砕して色素を抽出した結果です。スサビノリには紅色のフィコビリン、緑色のクロロフィル、黄色のカロチノイドなどの光合成色素が含まれています。陸上植物の葉は多量のクロロフィルと少量のカロチノイドが含むため,緑色に見えます。スサビノリにもクロロフィルやカロチノイドが含まれていますが,多量な色素はフィコビリンのために藻体は紅色を呈しています。これらの色素は葉緑体の中に含まれています。
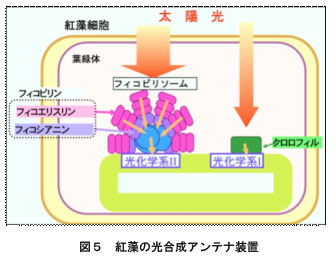 フィコビリンはおもにフィコシアニンやフィコエリスリンが含まれ,これらは光合成の反応に必要な光を集めるアンテナ装置(フィコビリソーム)の構成色素です(図5)。緑藻や陸上植物はフィコビリンをもたず、アンテナ色素としておもにクロロフィルを使っているため、葉などの光合成器官は緑色をしています。光合成を行うバクテリアの一種であるラン藻やスサビノリの属する紅藻はアンテナ色素としてフィコビリンを使う点で他の植物群とは異なっています。ただし,紅藻ではアンテナ装置としてフィコビリソームの他にクロロフィルでできたアンテナ装置(LHCI,図5の緑色の部分)をもち,ラン藻にはこの装置がない点は異なっています。「海苔の色落ち」は,葉緑体中のフィコビリソームが分解することにより起こることが明らかになっています。
フィコビリンはおもにフィコシアニンやフィコエリスリンが含まれ,これらは光合成の反応に必要な光を集めるアンテナ装置(フィコビリソーム)の構成色素です(図5)。緑藻や陸上植物はフィコビリンをもたず、アンテナ色素としておもにクロロフィルを使っているため、葉などの光合成器官は緑色をしています。光合成を行うバクテリアの一種であるラン藻やスサビノリの属する紅藻はアンテナ色素としてフィコビリンを使う点で他の植物群とは異なっています。ただし,紅藻ではアンテナ装置としてフィコビリソームの他にクロロフィルでできたアンテナ装置(LHCI,図5の緑色の部分)をもち,ラン藻にはこの装置がない点は異なっています。「海苔の色落ち」は,葉緑体中のフィコビリソームが分解することにより起こることが明らかになっています。
【5. ラン藻の「色落ち誘導遺伝子」】
ラン藻や紅藻は窒素やリンなど栄養分が欠乏するとフィコビリソームを分解し,色落ちを起こします。ラン藻では「色落ちを制御する遺伝子」としてNblAが同定されています。NblAと相同性のある遺伝子が紅藻の葉緑体にありますが,その働きは不明でした。私たちは,養殖ノリの色落ちの仕組みを考えるため,スサビノリを用いてこの遺伝子の発現様式について調べてきました。しかし,この遺伝子はラン藻とはまったく異なる発現様式を示しました。紅藻も葉緑体の祖先であるラン藻と同様に栄養欠乏によりフィコビリソームを分解しますが,その調節の仕組みはラン藻とは異なっているようでした。
【6. 赤潮による養殖ノリの色落ち】
外洋の海水中に含まれている栄養塩類の濃度は非常に薄く,栄養分の多くは陸地から供給されています。特に河川からの供給が大きな割合を占めています。海苔養殖が沿岸部で行われるのはこのような理由からです。植物プランクトンも,通常は栄養不足で増殖が制限されています。しかし,河川から大量の栄養分が供給されると,海藻よりも成長速度の大きい植物プランクトンが一時的に大量に増殖し,その結果,海水中の栄養塩類のうち特に窒素やリンの濃度が急激に低下します。栄養分がなくなると赤潮プランクトンも消滅しますが,海苔は海水中の栄養不足のため色落ちが起こります。すなわち,赤潮による海苔の色落ちはノリの栄養ストレスに対する応答と考えられています。
【7. 窒素欠乏と硫酸銅処理による色落ち】
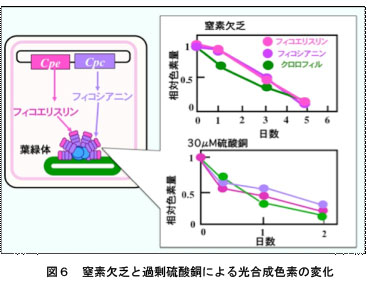 色落ちは栄養欠乏だけではなく,病原菌の感染や高水温,重金属添加などの色々なストレスによって起こります。これらの異なるストレス種により誘導される色落ち反応(フィコビリソーム分解反応)は同じメカニズムで起こるのでしょうか?一昨年から,窒素欠乏と硫酸銅ストレスに対するスサビノリの応答や色落ち過程で活性化する遺伝子の働きを調べてきました。図6のように,窒素欠乏でも硫酸銅処理でもフィコビリソームの分解は起こります。どちらの場合もフィコエリスリン,フィコシアニン,クロロフィルがほぼ一定の速度で減少していることから,どちらも同じメカニズムによって進行している可能性が考えられました。しかし,色落ち過程でみられる葉緑体の挙動やストレス処理条件から正常な条件に戻したときの細胞の応答には違いがみられました。通常状態では,葉緑体は細胞膜に足を伸ばした金平糖様の構造をしています。
色落ちは栄養欠乏だけではなく,病原菌の感染や高水温,重金属添加などの色々なストレスによって起こります。これらの異なるストレス種により誘導される色落ち反応(フィコビリソーム分解反応)は同じメカニズムで起こるのでしょうか?一昨年から,窒素欠乏と硫酸銅ストレスに対するスサビノリの応答や色落ち過程で活性化する遺伝子の働きを調べてきました。図6のように,窒素欠乏でも硫酸銅処理でもフィコビリソームの分解は起こります。どちらの場合もフィコエリスリン,フィコシアニン,クロロフィルがほぼ一定の速度で減少していることから,どちらも同じメカニズムによって進行している可能性が考えられました。しかし,色落ち過程でみられる葉緑体の挙動やストレス処理条件から正常な条件に戻したときの細胞の応答には違いがみられました。通常状態では,葉緑体は細胞膜に足を伸ばした金平糖様の構造をしています。
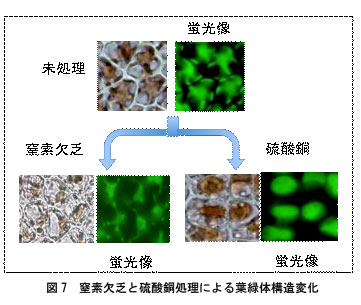 窒素欠乏では金平糖様の形態は保持したまま,葉緑体が収縮します(図7,窒素欠乏)。窒素欠乏5日目までの細胞であれば,正常培地に移して培養すると,成長が再開し,フィコビリン色素量や葉緑体の形態はもとに戻ります。一方,30μM銅処理細胞では,フィコビリン量が減少していない色落ちの初期段階の細胞では,葉緑体は金平糖様の構造から球形へと変化します(図7,硫酸銅)。さらに処理時間の経過とともに葉緑体が破壊され,細胞は死滅します。硫酸銅処理の場合,色落ち初期段階の細胞を硫酸銅を含まない培地に戻しても成長は再開しませんでした。
窒素欠乏では金平糖様の形態は保持したまま,葉緑体が収縮します(図7,窒素欠乏)。窒素欠乏5日目までの細胞であれば,正常培地に移して培養すると,成長が再開し,フィコビリン色素量や葉緑体の形態はもとに戻ります。一方,30μM銅処理細胞では,フィコビリン量が減少していない色落ちの初期段階の細胞では,葉緑体は金平糖様の構造から球形へと変化します(図7,硫酸銅)。さらに処理時間の経過とともに葉緑体が破壊され,細胞は死滅します。硫酸銅処理の場合,色落ち初期段階の細胞を硫酸銅を含まない培地に戻しても成長は再開しませんでした。
私達は,紅藻スサビノリがもつ「色落ちの仕組み」のうち少なくとも一部は,葉緑体の祖先であるラン藻から受け継がれていると予想し研究を進めきたが,残念ながら未だにこの考えを指示する証拠は得られていない。一方,スサビノリは真核生物であり,ラン藻とは異なるフィコビリソーム合成・分解システムを獲得している可能性があります。種子植物では栄養欠乏における細胞内成分の分解システムとしてオートファジーが知られています。また,病原菌に対する感染応答として,感染細胞が積極的に自殺する(プログラム細胞死)も知られています。上述のスサビノリの硫酸銅処理では,プログラム細胞死が誘発されている可能性も考えられます。スサビノリのESTデーターベースにもプログラム細胞死やオートファジー関連の遺伝子が登録されていることから,今後は,スサビノリの色落ちをこのような真核生物のストレス応答遺伝子の点からも解析を進める予定です。