
養殖ノリとアオサ類の遺伝子解析
熊本大学沿岸域環境科学教育研究センター
教授 瀧尾 進
平成22年11月4日(木)
はじめに
養殖ノリは有明海・八代海における重要な水産資源のひとつですが、平成12年には深刻な“色落ち”問題などで生産量は激減し、社会問題となりました。私は、平成13年の沿岸域環境科学教育研究センター設置を機にノリについて基礎研究を開始し、現在まで「スサビノリ(養殖ノリ)色落ちの分子機構」と「スサビノリのレトロトランスポゾン」を中心に研究を進めてきました。3年前から、逸見教授との共同研究として、干潟の環境保全に重要な塩生植物のうち、世界中に分布するハママツナの遺伝的多様性解析も始めました。さらに、各地で大量発生が問題になっているアオサについても逸見教授との共同研究として昨年から研究を始めました。ハママツナやアオサの研究は始まったばかりですが、今回はアオサの研究について紹介致します。
アオサの植物学上の特徴
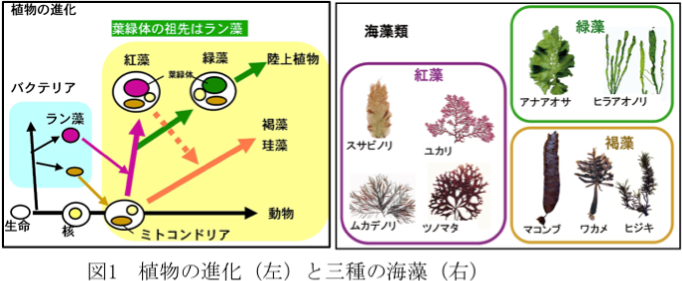
生物は原核生物(バクテリア)と真核生物の二つに大別できます(図1左、植物の進化)。原核生物では遺伝子の本体であるDNAが裸の状態で細胞内に存在していますが、真核生物ではDNAは核とよばれる膜に包まれた部位に閉じこめられています。真核生物はさらに植物と動物に大別することができ、植物ではDNAは核の他にミトコンドリアや葉緑体にも含まれています。
植物の進化において、光合成を行う原核生物であるラン藻が植物の祖先となる細胞に取り込まれて、やがて細胞内で共生し現在の葉緑体になったと考えられています(一次共生)。すなわち、紅藻は植物進化において最初に出現した植物と言えます。紅藻が出現した後、緑藻が生まれ、その仲間は陸上へと進出しました。スサビノリは紅藻で、アオサ類は緑藻に分類されます。
なお、コンブやワカメなどの褐藻類や赤潮プランクトンに含まれる珪藻や渦鞭毛藻類は、紅藻が動物細胞にもう一度取り込まれることによって生まれた(二次共生)植物と考えられています。
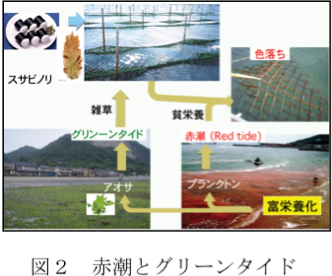
赤潮(red tide)とグリーンタイド
「海苔の色落ち」は、赤潮プランクトンの発生などにより海水中の栄養窒素濃度が低下し、海苔の色素が分解することにより起こることが明らかになっています。沿岸域の環境問題として赤潮(red tide)は良く知られていますが、最近ではアオサ類が大量発生するグリーンタイド(green tide)も大きな問題となっています(図2)。グリーンタイドは日本各地でみられ、九州においても博多湾などは良く知られています。一方、最近まで熊本県では他県ほど大きな問題とはなっていませんでした。しかし、一昨年菊池川河口で今までにない大量発生がみられ、今年も引き続き発生していることから、熊本県でも新たな環境問題として本格的に取り組む必要がでてきました。
スサビノリとアオサの生活環
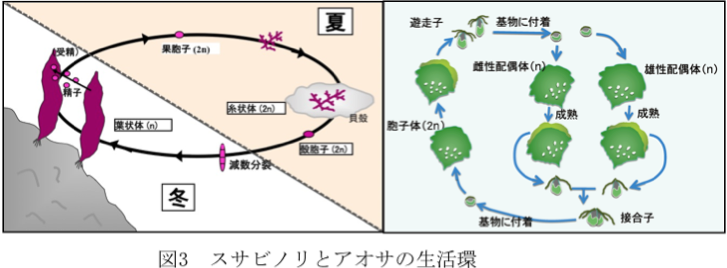
図3(左)はスサビノリの一生の模式図です。食用となる葉状体は半数世代(n、染色体数が2nの半分)で冬に生長します。春先になり精子と卵が葉状体に形成され、受精により受精卵(果胞子)ができ複数世代(2n)となります。果胞子が発芽すると葉状体とは全く異なる形態の糸状体となります。しかも、糸状体は貝殻の表面に付着し、その後、殻の中に侵入して殻の中でカビの菌糸のように増殖します。糸状体の生長は夏期に行われ、秋になると糸状体の一部から殻胞子(2n)が作られ、これが減数分裂した後に発芽し、半数世代の小さな葉状体となり、やがて大きな葉状体へと成長します。
図3(右)はアオサの一生の模式図です。アオサはスサビノリとは異なり、半数世代(n世代、配偶体)と倍数世代(2n世代、胞子体)が同じ形をしていて区別ができません。配偶体は雄と雌がありますが、これも形態上は区別できません。胞子体が成熟すると、藻体周辺から4本の鞭毛を有する遊走子が遊離します。遊走子が有性配偶体になり熟成すると、性別があり2本の鞭毛を有する配偶子が遊離します。雌雄の配偶子は接合して胞子体に成長しますが、配偶子がそのまま同性の配偶体になり無性的に増殖を繰り返すことができる種もあります。遊走子や配偶子を放出した成熟個体はやがて枯死します。しかし、理由は不明ですが、成熟能を失って無限に無性的に増殖することができるような不稔株も見つかっています。グリーンタイドとして大量発生するアオサ類にはこのような不稔性のアオサであることが多いです。
アオサの分類
アオサ科(Ulvaceae)は、 1.ヒメアオノリ属(Blidingia)、2.ヤブレグサ属(Umbraulva)、3.アオノリ属(Enteromorpha)、4.ペルクルサリア属(Percursaria)、5.アオサ属(Ulva)、6.クロヒトエグサ属(Ulvaria)のおもに6つ属からなっています。しかし、アオサ類は体制が単純で生育環境によって形態が大きく変異することがあり、野外から採取したサンプルの形態的特徴に加えて、実験室内での培養でもこのことが検証され、従来の分類は遺伝子配列の比較から、再検討されています。特に、アオノリ属は属名がEnteromophaとして、アオサ属のUlvaとは異なるグループとして扱われていましたが、最近では、アオノリ類はアオサ属に組み込まれています。
熊本県のアオサ類の遺伝子解析
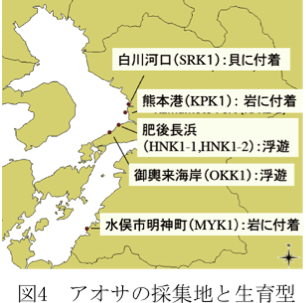 2009年の12月から2010年の4月にかけて熊本県内各地の海岸からアオサ類を採取し、遺伝子解析をおこないました(図4)。
2009年の12月から2010年の4月にかけて熊本県内各地の海岸からアオサ類を採取し、遺伝子解析をおこないました(図4)。
サンプルは液体窒素で破砕し、全DNAを抽出して遺伝子解析を行いました。残りの藻体の一部はスサビノリ培養と同じ培養液に入れ、継代培養を行いました。冬期に採集したサンプルなので、スサビノリと同じ培養条件(15℃、10時間明/14時間暗)で培養を行っています。肥後長浜から採取したサンプルは当初は1種類と考えて継代培養を行っていましたが、継代培養後に遺伝子解析を行ったところ、2種類のサンプルが混在していることが判明し、HNK1-1とHNK1-2と株名として独立して培養を行っています。
抽出した全DNAを用いて、核ゲノムにコードされるリボソーム遺伝子の2つのスペーサー領域(ITS1とITS2)をPCRにより増幅して、この領域の塩基配列を決定しました。
調べた6株のうち5株はアナアオサ(Ulva pertusa)と同様の配列を示しました。しかし、HNK1-1株はアナアオサとは明らかに異なり、ミナミアオサと類似するグループに近いことがわかりました。
大量発生するアオサ類はアナアオサと一般的には言われていますが、最近では新種のミナミアオサも含まれていることが明らかになっています。現在日本では、ミナミアオサのITS配列は愛知(三河湾)、高知(土佐)、福岡、沖縄(那覇市)から6クローンの配列が登録されおり、それらの配列は100%一致していました。
 しかし、HNK1-1とは一部異なっていました。系統樹で同じグループに含まれるリボンアオサ,アミアオサ、コツブアオサはそれぞれ一カ所からのサンプルしか登録されていませんが、HNK1-1株の配列はそれらいずれの配列とも一部異なっていました。HNK1-1と同じグループ含まれたリボンアオサ、ミナミアオサ、アミアオサ、コツブアオサは藻体の葉縁部に小さい鋸歯が見られる特徴があります。アナアオサにはこのような鋸歯はありません。HNK1-1株には鋸歯がみられます。一方、ITS配列からアナアオサと同定された他の5株はいずれも鋸歯は観察できませんでした(図5)。
しかし、HNK1-1とは一部異なっていました。系統樹で同じグループに含まれるリボンアオサ,アミアオサ、コツブアオサはそれぞれ一カ所からのサンプルしか登録されていませんが、HNK1-1株の配列はそれらいずれの配列とも一部異なっていました。HNK1-1と同じグループ含まれたリボンアオサ、ミナミアオサ、アミアオサ、コツブアオサは藻体の葉縁部に小さい鋸歯が見られる特徴があります。アナアオサにはこのような鋸歯はありません。HNK1-1株には鋸歯がみられます。一方、ITS配列からアナアオサと同定された他の5株はいずれも鋸歯は観察できませんでした(図5)。
HNK1-1株は培養開始後数ヶ月は成熟することなく増殖していましたので、不稔性であると推定していました。ただし、ミナミアオサはアナアオサに比べると成熟が遅いことが報告されています。したがって、HNK1-1株が本当に不稔性であるかについては培養温度や光条件を変えて検討する必要があるでしょう。
おわりに
熊本県のアオサの遺伝子解析は始まったばかりですが、今回の研究により大量発生の原因となる種がアナアオサだけではなくミナミアオサに近縁の種も存在する可能性が示唆されました。福岡で検出されるミナミアオサが熊本にも拡散していることが予想されましたが、 HNK1-1株は従来のミナミアオサとは異なっているようです。では、従来のミナミアオサは熊本にはいないのでしょうか?HNK1-1はどこからやってきたのでしょうか?今回用いた材料は、とくにアオサが大量発生している海岸から採取したものではありません。しかし、HNK1-1はサンプル採取時には比較的大量に発生していましたが、一ヶ月後にはほとんど消失していました。今後は、大量発生の起こっている場所からのサンプルについて調査する必要があります。